


成纤维细胞生长因子受体2b(FGFR2b):结构、信号传导和生物学功能
成纤维细胞生长因子受体2b(FGFR2b)是成纤维细胞生长因子受体2(FGFR2)的亚型之一,属于受体酪氨酸激酶(RTK)家族。FGFR2b与其他FGFR家族成员一样,在细胞膜上表达,并通过其独特的结构成分和信号传导通路在多种细胞过程中发挥关键作用。本文深入分析了FGFR2b的结构、配体诱导的信号传导机制及其生物学功能,强调其在发育、组织修复和疾病发病机制中的重要性。

- 最新进展
- 产品信息
最新进展
摘要
成纤维细胞生长因子受体2b(FGFR2b)是成纤维细胞生长因子受体2(FGFR2)的亚型之一,属于受体酪氨酸激酶(RTK)家族。FGFR2b与其他FGFR家族成员一样,在细胞膜上表达,并通过其独特的结构成分和信号传导通路在多种细胞过程中发挥关键作用。本文深入分析了FGFR2b的结构、配体诱导的信号传导机制及其生物学功能,强调其在发育、组织修复和疾病发病机制中的重要性。成纤维细胞生长因子(FGF)信号传导通路对于胚胎发育、组织修复、细胞增殖、分化和存活等多种生物学过程至关重要。FGF信号传导通路由成纤维细胞生长因子受体(FGFR)家族介导,这些受体是受体酪氨酸激酶(RTK)。FGFR能够结合FGF配体,激活下游信号传导通路,从而调节细胞行为。在FGFR家族成员中,FGFR2b因其独特的亚型特异性功能以及在多种生理和病理过程中的参与而备受关注。
2. FGFR2b的结构
FGFR2b是通过FGFR2基因的可变剪接产生的亚型之一。与FGFR家族的其他成员类似,FGFR2b是一种跨膜受体酪氨酸激酶,由三个主要结构元件组成:
2.1 细胞外结构域
FGFR2b的细胞外结构域包含三个免疫球蛋白样亚结构域(D1、D2和D3)。这些亚结构域对于配体结合至关重要,特别是与成纤维细胞生长因子(FGF)配体的结合。其中,D3亚结构域对于与FGF配体的高亲和力结合尤为重要,而D1和D2亚结构域则有助于配体结合的特异性和亲和力。FGFR2b与FGF配体之间的相互作用通常由肝素硫酸蛋白多糖(HSPGs)协助,它们作为共受体,增强配体-受体复合物的结合亲和力和稳定性。
2.2 跨膜螺旋
跨膜螺旋是一个单次跨膜结构,将受体锚定在细胞膜中。该结构域将细胞外结构域与细胞内结构域连接起来,确保受体在质膜中的正确定位和稳定。跨膜螺旋对于维持受体的结构完整性以及其作为信号传导分子的功能至关重要。
2.3 细胞内酪氨酸激酶结构域
细胞内酪氨酸激酶结构域是受体的催化核心,在信号传导中发挥关键作用。配体结合后,受体发生构象变化,导致FGFR2b分子的二聚化。这种二聚化使细胞内激酶结构域相互靠近,从而允许激酶结构域内特定酪氨酸残基的自身磷酸化。磷酸化的酪氨酸残基作为下游信号传导分子的停靠位点,启动一系列细胞内信号传导事件。
3. FGFR2b的信号传导机制
FGFR2b信号传导通路的激活始于FGF配体与受体细胞外结构域的结合。这种结合事件诱导受体发生构象变化,导致受体二聚化,随后激活细胞内酪氨酸激酶结构域。激活的激酶结构域磷酸化特定的酪氨酸残基,为下游信号传导分子创造结合位点。FGFR2b激活的主要信号传导通路包括:
3.1 RAS-MAPK通路
RAS-MAPK通路是FGFR2b激活的最显著的信号传导级联反应之一。受体激活后,磷酸化的酪氨酸残基招募适配蛋白,如FGFR底物2(FRS2)、SOS(七号基因之子)和生长因子受体结合蛋白2(GRB2)。这些蛋白促进RAS的激活,进而激活一系列下游激酶,包括RAF、MEK和ERK(细胞外信号调节激酶)。ERK的激活导致多种转录因子的磷酸化,最终导致基因表达的变化,从而促进细胞增殖、分化和存活。
3.2 PI3K-mTOR-AKT通路
PI3K-mTOR-AKT通路是FGFR2b激活的另一个关键信号传导通路。该通路的激活始于磷脂酰肌醇3激酶(PI3K)被招募到受体上的磷酸化酪氨酸残基。PI3K催化产生磷脂酰肌醇-3,4,5-三磷酸(PIP3),进而招募并激活蛋白激酶B(AKT)。AKT随后磷酸化并激活雷帕霉素靶蛋白(mTOR),这是细胞生长、代谢和存活的关键调节因子。PI3K-mTOR-AKT通路对于促进细胞存活、蛋白质合成和代谢调节至关重要。
3.3 STAT3通路
信号转导与转录激活因子3(STAT3)通路也由FGFR2b激活。受体激活后,STAT3被招募到磷酸化的酪氨酸残基上并发生磷酸化。磷酸化的STAT3随后二聚化并转移到细胞核中,在那里结合特定的DNA序列,调节涉及细胞增殖、存活和免疫反应的基因表达。
4. FGFR2b的生物学功能
4.1 胚胎发育
FGFR2b在胚胎发育中发挥着关键作用,特别是在多种组织和器官的形成和分化过程中。在胚胎发生过程中,FGFR2b介导的信号传导对于上皮组织的正常发育至关重要,包括皮肤、胃肠道和呼吸系统的发育。FGF配体与FGFR2b的结合促进了上皮细胞的增殖和分化,确保了这些组织的正确形成和功能。
4.2 组织修复和再生
FGFR2b还参与组织损伤后的修复和再生。在组织受损后,FGF配体被释放并与受损区域细胞表面的FGFR2b结合。这种结合激活了受体并启动了促进细胞增殖、迁移和分化的信号传导通路,从而形成新组织并恢复组织功能。例如,在伤口愈合过程中,FGFR2b信号传导对于皮肤的再上皮化和肉芽组织的形成至关重要。
4.3 疾病发病机制
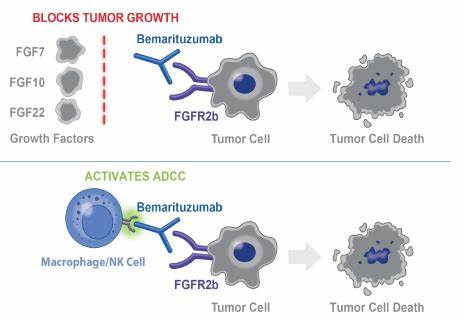
FGFR2b信号传导的异常激活与多种疾病的发生有关,特别是在癌症的背景下。FGFR2b的突变或过表达可能导致下游信号传导通路的持续激活,从而促进细胞不受控制的增殖、存活和迁移。这种FGFR2b信号传导的失调已在多种癌症中观察到,包括胃癌、乳腺癌和肺癌,它们促进了肿瘤的生长、侵袭和转移。此外,FGFR2b信号传导还与其他病理状况有关,如纤维化和心血管疾病,在这些疾病中,它可能有助于组织重塑和炎症。
5. 治疗意义
鉴于FGFR2b在多种生理和病理过程中的关键作用,靶向FGFR2b信号传导已成为多种疾病的有前途的治疗策略。在癌症中,FGFR2b抑制剂已在临床前和临床研究中显示出潜力,能够抑制肿瘤生长和转移。这些抑制剂包括小分子酪氨酸激酶抑制剂和单克隆抗体,它们特异性地靶向FGFR2b,阻断其激活和下游信号传导。除了癌症外,靶向FGFR2b信号传导还可能为其他以异常组织修复和纤维化为特征的疾病提供治疗益处,如慢性肾脏病和特发性肺纤维化。
6. 结论
成纤维细胞生长因子受体2b(FGFR2b)是FGFR家族的关键成员,在胚胎发育、组织修复和疾病发病机制中发挥着关键作用。其独特的结构特征和信号传导机制使其能够通过激活下游信号传导通路(如RAS-MAPK、PI3K-mTOR-AKT和STAT3)调节广泛的生物学过程。了解FGFR2b的结构和功能为其在正常生理和疾病中的作用提供了宝贵的见解,并强调了其作为多种疾病治疗靶点的潜力。未来的研究将继续阐明FGFR2b信号传导的复杂机制,并探索其在临床环境中的治疗应用。
产品信息
杭州斯达特 (www.starter-bio.com)志在为全球生命科学行业提供优质的抗体、蛋白、试剂盒等产品及研发服务。依托多个开发平台:重组兔单抗、重组鼠单抗、快速鼠单抗、重组蛋白开发平台(E.coli,CHO,HEK293,InsectCells),已正式通过欧盟98/79/EC认证、ISO9001认证、ISO13485。
 查看更多产品
查看更多产品